
On appelle "perception" ce que notre œil capte inconsciemment. Mais s’il y a une volonté de regarder un objet précis, on parle alors "d’attention visuelle". Derrière ce regard dirigé par la pensée, il y a tout un réseau de neurones qui décompose le travail en microtâches pour ensuite en reconstruire le sens.
 De la neurophysiologie cognitive
De la neurophysiologie cognitive
Derrière un simple regard, notre cerveau met en branle une mécanique complexe, mais efficace, qui sollicite des milliards de neurones. Le tout suit un ordre de marche assez bien huilé.
Les mécanismes cérébraux de la perception des différents éléments d’une scène visuelle – par exemple : forme, couleur et mouvement – sont assez bien connus. Aussi de nombreux chercheurs s’attardent-ils maintenant aux processus cognitifs volontaires qui modulent le comportement du cortex visuel.
C’est le cas de notre Laboratoire de neurophysiologie cognitive, qui, sous la tutelle du Dr Julio Martinez-Trujillo, s’intéresse à l’attention visuelle. On appelle « perception » ce que notre œil capte inconsciemment. Mais s’il y a une volonté de regarder un objet précis, on parle alors d’« attention visuelle ».
Que l’on fixe intensément ou que l’on regarde discrètement du coin de l’œil, l’attention visuelle est souvent décrite comme un projecteur accentuant certaines caractéristiques du stimulus (contraste, couleur, etc.), tout en filtrant ce qui pourrait distraire. Cette sélection facilite le traitement par le cortex visuel.
L’attention multifocale
Si l’on fixe un chat dormant sur une chaise, le traitement se fait simplement. Mais que se passe-t-il si l’on doit activement suivre plusieurs « objets » en mouvement dans une même scène? Prenons, par exemple, cette partie de football où le quart-arrière (no 6) porte son attention sur le receveur de passe (no 21), tout en restant attentif au joueur de la ligne défensive adverse qui le menace d’un placage (en gris, les éléments hors de la zone d'attention visuelle).
Plusieurs hypothèses ont été émises pour expliquer comment le cerveau humain gère une telle situation où de multiples objets nécessitent une attention soutenue. Il a d'abord été proposé que le focus d’attention, pour reprendre l’analogie du projecteur, s’élargit afin d’illuminer tous les éléments observés, y compris les éléments non pertinents présents entre ceux-ci1. La deuxième hypothèse stipule que le focus de l’attention se déplace rapidement d’un élément à un autre et qu’un seul objet en est la cible à chaque instant2. La troisième hypothèse avance qu’il est possible de maintenir plusieurs foyers d’attention simultanément3. Celle-ci a été récemment démontrée empiriquement dans le laboratoire du Dr. Julio Martinez-Trujillo de l’Université McGill4.

Les chemins de l’information visuelle
Afin de pouvoir quantifier un concept aussi abstrait que l’attention visuelle, il est primordial de comprendre les mécanismes cérébraux de la vision. En effet, puisque la reproductibilité est à la base de toute science, il faut fournir des mesures concrètes sur lesquelles appuyer chacune de nos théories. Savoir quoi mesurer, quand le faire et surtout où prendre les mesures représente des concepts clefs en neuroscience. Avec un organe aussi complexe que le cerveau, chaque procédure doit être planifiée avec la plus grande précision. Voyons alors les bases du traitement visuel par le cortex afin de répondre à ces questions.
Au niveau du système visuel, il existe deux voies de traitement fonctionnellement spécialisées. Une voie « ventrale » impliquée dans l’identification et la reconnaissance de formes et des couleurs, et une voie « dorsale » impliquée dans la perception du mouvement et la localisation spatiale.
Ces deux voies traversent plusieurs régions corticales et ce, dans un ordre hiérarchique et séquentiel. Cela signifie simplement que l’information visuelle, en provenance de la rétine, entre dans la cascade par le cortex visuel primaire, communément appelé V1. Ce dernier la convoie vers plusieurs régions hautement spécialisées qui séparent l'information en ses constituants primaires, tels la couleur, l’orientation des segments composants une forme et la direction du mouvement.
Prenons par exemple la région V4, de la voie «ventrale», qui encode la couleur et certaines formes géométriques très simples, ou la région V5, de la voie «dorsale», qui encode la direction d’un mouvement linéaire. Cela signifie étonnamment qu’une personne ayant une lésion dans l’une ou l’autre de ces régions serait « aveugle » soit aux couleurs (achromatopsie), soit au mouvement (akinetopsie), tout en percevant parfaitement le reste du monde!
Certaines régions corticales intègrent l’information des formes géométriques simples et d'autres sont entièrement dévouées au traitement des visages.
Par la suite, l’information provenant de ces régions converge vers des régions dites supérieures où elle acquiert une complexité supplémentaire. Pour reprendre nos exemples, cela signifie que certaines régions intègrent l’information des formes géométriques simples provenant de V4 et encodent des stimuli beaucoup plus complexes, comme par exemple les visages! En effet, certaines régions corticales sont entièrement dévouées au traitement des visages. Il en est de même avec une région particulière recevant l’information provenant de V5, qui combine les différents mouvements linéaires et encode les mouvements de spirale, d’expansion et de contraction. Plus l’on avance dans la hiérarchie, plus les pièces du casse-tête s’imbriquent et plus la représentation mentale de la scène visuelle se complète.
Passons maintenant du plus grand vers le plus petit. Puisque nous sommes dans le cerveau, l’unité fonctionnelle à la base de toute action est bien sûr le neurone. Les neurones composant chacune de ces régions répondent bien évidemment aux stimuli spécifiques de leur région, mais seulement lorsque celui-ci est situé à un endroit bien précis du champ visuel.
Ce champ de vision neuronal, que nous appelons champ récepteur (CR), est spécifique mais non exclusif à chaque neurone. La totalité des CRs des neurones d’une région couvrent donc la totalité du champ visuel. Mentionnons également que, telle la complexité du stimulus encodé, la taille du champ récepteur des neurones s’élargit au fur et à mesure que l’on avance à travers la hiérarchie corticale. Puisque les régions supérieures de la hiérarchie intègrent l’information provenant des neurones des régions précédentes, par le fait même, elles fusionnent leurs champs récepteurs.
Au coeur de l'expérience, la mesure...
Sachant le rôle et la localisation de chaque région, il est désormais possible de construire notre expérience.
Quoi mesurer? L’activité de neurones individuels, à l’aide d’enregistrements électrophysiologiques; on voit le résultat dans la figure suivante. Les potentiels d’action, ou « décharges », de neurones individuels sont enregistrés à l’aide d’électrodes implantées dans le cortex. L’activité d’un neurone est directement quantifiée selon sa fréquence de décharge. Plus elle est élevée, plus un neurone est actif. Il est alors aisé de remarquer que le neurone de l’exemple réagit à la présentation du stimulus car son activité augmente fortement pendant cette période.
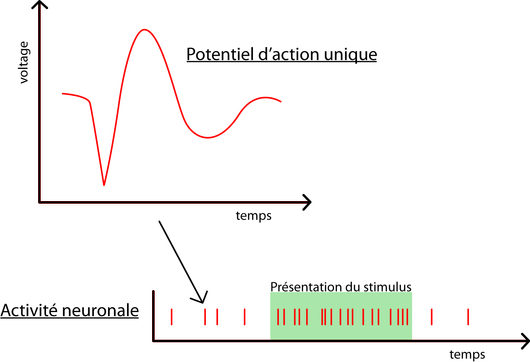
Quand mesurer? Lorsque le sujet est impliqué dans une tâche où il doit diviser son attention visuelle entre plusieurs cibles et ignorer les distracteurs.
Où mesurer? Puisque l’organisation est hiérarchique, un même stimulus active plusieurs régions successivement; il faut donc choisir la région spécialisée pour un stimulus donné, dont la taille sera ajustée pour correspondre approximativement au CR des neurones de cette région.
Stimuler un neurone à la fois
Notre objet d’étude est ici une population de neurones de la région médiane temporale (MT) du cortex visuel du macaque, correspondant à l’aire V5 chez l’humain. Ces neurones font partie du parcours dorsal et répondent aux mouvements linéaires. Tel que mentionné précédemment, les neurones de cette région sont impliqués dans la perception du mouvement, en répondant de manière sélective à une certaine direction et vitesse de mouvement. Le stimulus optimal pour « déclencher » la réponse d’un neurone de cette population pourrait donc être tout objet, dans notre cas un patron de points aléatoire se déplaçant vers la gauche et présenté à l’intérieur du CR. Un stimulus se déplaçant vers la droite ou positionné hors du CR n’engendreraient qu’une réponse très faible ou nulle.
La figure suivante illustre le stimulus optimal pouvant activer ce neurone. Le cercle gris (CR) et la flèche blanche (direction du mouvement) ne seraient bien sûr pas affichés. Le point blanc central représente simplement le centre de l’espace visuel et le point de fixation.

Nos neurones encodent donc des caractéristiques précises du stimulus. Rappelons que s’il y a de plus une « attention visuelle » sur celui-ci, la réponse sera accrue, comparativement à un même stimulus non ciblé. Il est primordial de garder ce concept en mémoire car toute l'expérience en découle.
Maintenant que nous savons comment activer nos neurones, il ne reste plus qu’à mesurer leur activité lorsque le stimulus présenté dans leur CR est pertinent ou non.
Ces neurones ne sont pas dans une boîte de Petri, mais in vivo dans le cerveau de macaques sur lesquels nous effectuons des enregistrements électrophysiologiques corticaux. L’animal est préalablement entraîné à répondre à un changement de direction du mouvement des patrons de points, ou à un changement de luminance du point de fixation central.
L’expérience fait appel à trois situations, illustrées dans la figure suivante :

1. Dans la première situation, l’animal doit détecter un changement dans la direction de mouvement de l’un ou l’autre des stimuli périphériques (représentés par les flèches rouges) cernant un stimulus similaire localisé à l’intérieur du CR (cercle blanc) du neurone dont on enregistre l’activité . Il est donc assumé que l’animal dirige son attention simultanément aux deux stimuli périphériques, et que le stimulus central à l’intérieur du CR agit à titre de distracteur.
2. Dans la seconde situation, l’animal doit détecter le changement dans le stimulus situé dans le CR du neurone (flèche blanche). L’attention de l’animal est ainsi dirigée sur ce stimulus, qui agit à titre de cible. Les deux autres stimuli sont donc des distracteurs.
3. Pour la troisième et dernière situation, l’animal doit porter son attention au point de fixation central et détecter un changement dans son intensité lumineuse. Tout autre stimulus est considéré comme distracteur.
Validation des trois hypothèses
Ces expériences permettent de valider les trois hypothèses précédemment présentées concernant l’allocation de l’attention.
Dans un premier cas, si le focus d’attention s’agrandit pour englober les deux stimuli pertinents, il engloberait du même coup le distracteur contenu dans le CR du neurone. Une augmentation de la réponse du neurone serait conséquemment observée si le stimulus du CR est cerné des deux autres stimuli recevant l’attention visuelle (situation 1), comparativement à la situation 3 (tous ignorés). Ce qui n’est pas le cas. Non seulement la fréquence de décharge du neurone ne croit pas, mais elle diminue, comme si le cerveau supprimait la réponse au distracteur. La 1ère hypothèse est donc invalidée.
Si le foyer d’attention se déplace rapidement d’une cible à l’autre, comme le voudrait la 2e hypothèse, nous devrions retrouver une distribution bimodale des temps de réaction et de la performance de l’animal durant la première situation comportementale. Nous aurions un temps plus court et un plus haut taux de succès lorsque le changement de direction s’effectue sur le stimulus recevant l’attention. De l’autre côté, nous aurions un temps de réaction plus long, ainsi qu’un plus haut taux d’erreur, lorsque le changement s’effectue sur le stimulus ignoré. À nouveau, ce phénomène n’a pas été observé.
Les temps de réaction présentent une distribution uni-modale et aucune différence dans la performance n’a été observée. Voilà un bon point pour la 3e hypothèse. La combinaison de ces résultats, et de quelques autres trop lourds à détailler, nous permet aisément d’affirmer qu’il est possible de maintenir plusieurs foyers d’attention simultanément. Malgré le fait que cette découverte fait grandement progresser notre connaissance sur l’attention visuelle, elle demeure un concept très complexe. L’attention multifocale n’en est certainement pas le seul mécanisme et nombreux sont ceux qui doivent toujours être expliqués voire découverts.
De fait, l’attention visuelle est un concept très difficilement mesurable. Pour l’étudier nous devons entraîner l’animal à la placer à un endroit spécifique sur demande, ce qui est extrêmement laborieux. De plus, il serait tout à fait possible qu’un mécanisme d’attention inverse existe, c’est-à-dire un mécanisme qui inhiberait activement les réponses neuronales à un stimulus volontairement ignoré. La résolution de ces problèmes nécessitera une panoplie d’expériences supplémentaires.
Un petit exercice pour tester votre attention
La vidéo suivante est un excellent exemple d’attention implicite et multifocale. Le visionnement en mode plein écran est conseillé.

Elle illustre non seulement qu’il est possible de maintenir plusieurs foyers d’attention, mais que les réponses neuronales aux distractions sont, tel que nous l’avons mentionné précédemment, inhibées, aussi évidentes soient-elles. Ce phénomène de l’attention visuelle est connu depuis plusieurs années, mais il est intéressant de maintenant pouvoir le mesurer sur le plan neuronal. Cette nouvelle perspective pourrait nous éclairer davantage sur les dynamiques des réseaux neuronaux impliqués dans l’attribution de l’attention, qu’elle soit visuelle ou non, afin de mieux comprendre certaines conditions tels les déficits d’attention et l’autisme.
- 1ERIKSEN, C.W. et J.D. ST. JAMES (1986). « Visual attention within and around the field of focal attention: a zoom lens model », Perception & Psychophysics, 40, p. 225-240.
- 2POSNER, M.I., C.R. SNYDER et B.J. DAVIDSON (1980). « Attention and the detection of signals », Journal of Experimental Psychology, 109, p. 160-174.
- 3CASTIELLO, U. et C. UMILTÀ (1992). « Splitting focal attention », Journal of Experimental Psychology. Hum. Percept. Perform., 18, p. 837-848.
- 4NIEBERGALL, R., P.S. KHAYAT, S.TREUE et J.C. MARTINEZ-TRUJILLO (2011).« Multifocal attention filters targets from distracters within and beyond primate MT neurons’ receptive field boundaries », Neuron, 72, p. 1067-1079.
- Guillaume Doucet
Université McGill
Guillaume Doucet est étudiant au doctorat en physiologie à l’Université McGill (Montréal). Il étudie les mécanismes neuronaux du contrôle de l’attention visuelle par le cortex préfrontal, et plus précisément le biais attentionnel envers les stimuli socialement importants (visages et émotions) versus les stimuli non sociaux (objets inanimés), et ce chez les primates (humains et non-humains).
Vous aimez cet article?
Soutenez l’importance de la recherche en devenant membre de l’Acfas.
Devenir membre


